“Ah, memoria, nemica mortale del mio riposo!”
— Miguel de Cervantes
La corteccia entorinale laterale svolge un ruolo cruciale nella formazione e richiamo dei ricordi nel cervello umano. Questa regione del cervello è coinvolta nel processo di memorizzazione delle informazioni e nella creazione di connessioni sinaptiche che permettono il recupero dei ricordi. La comprensione di questi meccanismi è fondamentale per la comprensione delle malattie neurodegenerative, come l’Alzheimer, che colpiscono proprio questa area cerebrale. Recentemente, sono state sviluppate nuove metodologie di ricerca per indagare ulteriormente sulla funzione della corteccia entorinale laterale. Questo articolo esplorerà il ruolo di questa regione cerebrale nella memoria e le prospettive future nella neuroscienza.
Tozzi, F., Guglielmo, S., Paraciani, C., van den Oever, M. C., Mainardi, M., Cattaneo, A., & Origlia, N. (2024). Involvement of a lateral entorhinal cortex engram in episodic-like memory recall. Cell Reports, 43(10). doi:10.1016/j.celrep.2024.114795
Il ruolo cruciale della corteccia entorinale nella memoria
La corteccia entorinale, situata nella parte mediale del lobo temporale, svolge un ruolo cruciale nella memoria. Questa regione del cervello è coinvolta nella formazione e nel richiamo dei ricordi, giocando un ruolo fondamentale nel processo di apprendimento e nella navigazione spaziale. La corteccia entorinale riceve input da diverse aree cerebrali, tra cui l’ippocampo, ed è responsabile della codifica e dell’integrazione delle informazioni sensoriali e cognitive. Attraverso le sue connessioni con altre regioni cerebrali, la corteccia entorinale contribuisce alla creazione di reti neurali complesse che sostengono la memoria a lungo termine. Studi recenti hanno evidenziato l’importanza di questa regione nella memoria episodica, nella memoria spaziale e nel riconoscimento degli oggetti. Comprendere il ruolo della corteccia entorinale nella memoria può avere importanti implicazioni per la comprensione e il trattamento delle malattie neurodegenerative che colpiscono questa regione del cervello.
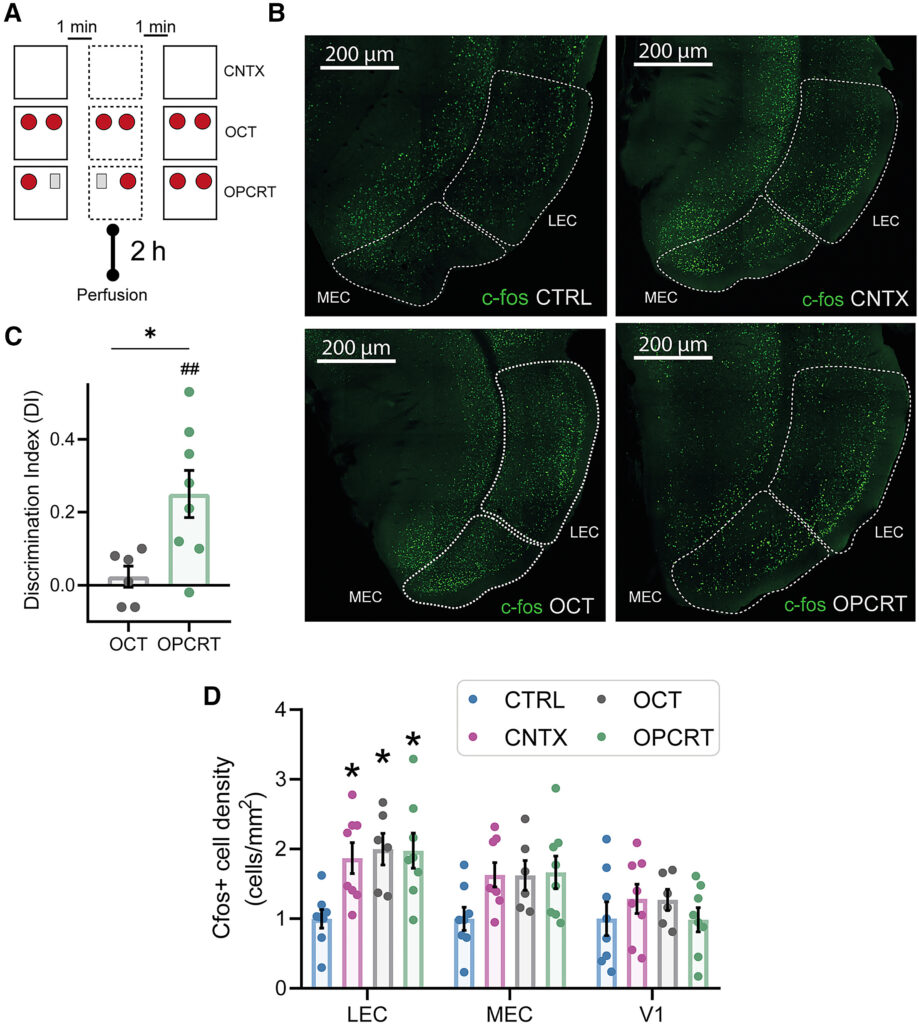
(A) Rappresentazione schematica dell'esposizione al contesto senza oggetti (gruppo CNTX), senza novità (esplorazione di due oggetti identici, OCT) e del test OPCRT, con perfusione degli animali eseguita 2 ore dopo il compito comportamentale.
(B) Immagini rappresentative che mostrano l'espressione di c-Fos (verde) nel LEC mediale e nel LEC nei vari gruppi sperimentali.
(C) L'indice di discriminazione (DI) dei topi sottoposti a OPCRT era significativamente più alto del caso (0,25 ± 0,06, n = 8, ##p = 0,0062, df = 7, t = 3,87, one-sample t test; p < 0,01), ma non è stata osservata alcuna differenza significativa dal livello casuale per il gruppo OCT (0,02 ± 0,03, n = 6, p = 0,46, df = 5, t = 0,80, one-sample t test). È stata osservata una differenza significativa tra il gruppo OCT e OPCRT (0,02 ± 0,03, n = 6 OCT vs. 0,25 ± 0,06, n = 8 OPCRT, df = 12, t = 2,85, ∗p = 0,015 test t a campione).
(D) È stata osservata una differenza significativa nella densità delle cellule c-Fos+ nel LEC (LEC) (ANOVA a due vie con test di confronto multiplo di Tukey: CTRL 1 ± 0,13 n = 8 vs. OPCRT 1,99 ± 0,22 n = 8, ∗p = 0,0127; CTRL vs. CNTX 1,87 ± 0,22 n = 8, ∗p = 0,0152; CTRL vs. OCT 1,97 ± 0,25 n = 6, ∗p = 0,0155). Non sono state osservate differenze significative nella MEC (CTRL 1 ± 0,16 n = 8 topi vs. OPCRT 1,66 ± 0,23 n = 8 topi, p = 0,095; CTRL vs. CNTX 1,63 ± 0,2 n = 8 topi, p = 0,119; CTRL vs. OCT 1,62 ± 0,21, p = 0,177) e nella corteccia visiva primaria (V1) (ANOVA a due vie con test di confronto multiplo di Tukey: CTRL 1 ± 0,24 n = 8 topi vs. OPCRT 0,985 ± 0,17 n = 8 topi, p = 0,99; CTRL vs. OCT 1,27 ± 0,15 n = 6 topi, p = 0,80; CTRL vs. CNTX 1,28 ± 0,21 n = 8 topi, p = 0,73). Valori espressi come cellule c-Fos+/mm2 rispetto a CTRL. I dati sono riportati come media ± SEM. Barra di scala in (B), 20 μm.
Meccanismi di formazione e richiamo dei ricordi
I meccanismi di formazione e richiamo dei ricordi rappresentano un aspetto fondamentale nel funzionamento della memoria. Durante il processo di formazione dei ricordi, le informazioni vengono elaborate e codificate nella corteccia entorinale laterale, una regione del cervello coinvolta nella memoria episodica. Qui, le informazioni vengono trasformate in tracce mnemoniche attraverso complesse interazioni tra neuroni e sinapsi. Durante il richiamo dei ricordi, le tracce mnemoniche vengono riattivate, consentendo la rievocazione delle informazioni precedentemente memorizzate. Questo processo è influenzato da numerosi fattori, tra cui l’attenzione, l’emozione e l’associazione con altri ricordi. La comprensione dei meccanismi di formazione e richiamo dei ricordi è di vitale importanza per comprendere come funziona la memoria e per sviluppare nuove strategie per migliorare la memoria nelle malattie neurodegenerative. Ricerche recenti hanno utilizzato diverse metodologie, come l’elettrofisiologia e l’imaging cerebrale, per indagare questi processi e hanno fornito nuove prospettive sulle basi neurali della memoria.
Implicazioni per le malattie neurodegenerative
Le implicazioni per le malattie neurodegenerative legate alla corteccia entorinale sono di grande importanza nella ricerca scientifica. Questa regione del cervello è coinvolta nel processo di formazione e richiamo dei ricordi, e alterazioni a livello della corteccia entorinale possono portare a disturbi della memoria, come nel caso della malattia di Alzheimer. Studi recenti hanno evidenziato una correlazione tra la degenerazione della corteccia entorinale e l’insorgenza di queste malattie neurodegenerative. La comprensione dei meccanismi che portano alla perdita di funzionalità della corteccia entorinale potrebbe fornire importanti indicazioni per lo sviluppo di nuove terapie e trattamenti. Inoltre, l’identificazione di biomarcatori specifici associati alle malattie neurodegenerative potrebbe consentire una diagnosi precoce e un intervento tempestivo per rallentare o prevenire la progressione delle patologie. Le implicazioni per le malattie neurodegenerative nella corteccia entorinale rappresentano un campo di ricerca in continua evoluzione e promettente per il futuro della neuroscienza.
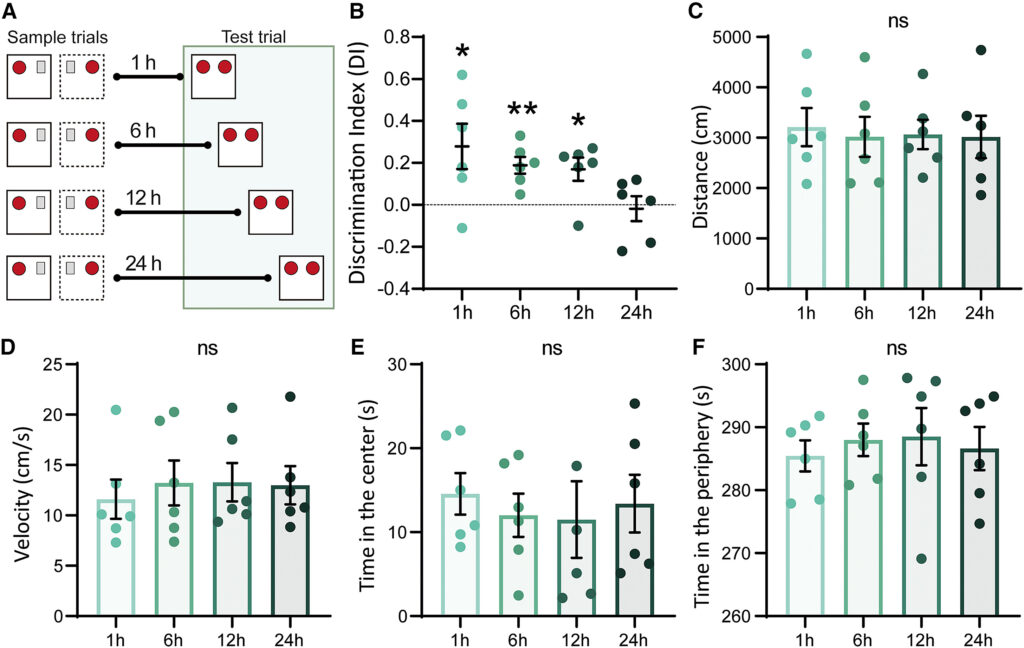
(A) Rappresentazione schematica del test OPCRT ritardato, con la prova condotta a diversi intervalli di tempo (1, 6, 12 o 24 ore) dopo la presentazione della prova campione.
(B) La DIs per l'associazione con il nuovo OPC mostra un calo significativo con l'aumentare degli intervalli di tempo dalla presentazione della prova campione (p = 0,037, test di Kruskal-Wallis). I topi sono stati in grado di richiamare la memoria simil-episodica precedentemente acquisita a 1 h (0,28 ± 0,11, n = 6, p = 0,049, df = 5, t = 2,58), 6 h (0,19 ± 0,04, n = 6, p = 0,0053, df = 5, t = 4,71), o 12 h (0,19 ± 0,04, n = 6, df = 5, t = 2,58). 71), o 12 h (0,17 ± 0,06, n = 6, p = 0,03, df = 5, t = 3,06; one-sample t test), mentre non è stata osservata alcuna memoria a 24 h (-0,018 ± 0,06, n = 6, p = 0,77, df = 5, t = 0,31, one-sample t test).
(C) Non è stata osservata alcuna differenza significativa nella distanza totale percorsa durante il test in campo aperto tra i gruppi sperimentali (n = 6, p = 0,97, F = 0,06, ANOVA a una via).
(D) La velocità dell'animale durante il test in campo aperto non differiva significativamente tra i gruppi (n = 6, p = 0,92, F = 0,16, ANOVA a una via).
(E) Il tempo trascorso dagli animali al centro dell'arena era simile tra i diversi gruppi sperimentali (n = 6, p = 0,92, f = 0,17, ANOVA a una via).
(F) Gli animali hanno trascorso una quantità di tempo comparabile nella periferia dell'arena (n = 6, p = 0,92, f = 0,17, ANOVA a una via). ∗p < 0,05, ∗∗p < 0,01. I dati sono riportati come media ± SEM.
Ricerche recenti e metodologie utilizzate
Negli ultimi anni, sono state condotte numerose ricerche sulla corteccia entorinale al fine di comprendere meglio il suo ruolo nella formazione e nel richiamo dei ricordi. Le metodologie utilizzate in queste ricerche sono state varie e hanno contribuito a fornire importanti informazioni sulla funzione di questa regione cerebrale. Tra le metodologie più utilizzate vi sono la microscopia a due fotoni, che consente di osservare l’attività neuronale in vivo, e l’uso di modelli animali come i topi transgenici. Inoltre, sono state sviluppate tecniche innovative come l’optogenetica, che permette di controllare selettivamente l’attività dei neuroni nella corteccia entorinale. Queste ricerche hanno dimostrato che la corteccia entorinale laterale svolge un ruolo cruciale nella codifica e nel recupero dei ricordi spaziali. Comprendere le metodologie utilizzate in queste ricerche è fondamentale per avanzare nella nostra conoscenza della memoria e delle malattie neurodegenerative associate ad essa.
Prospettive future nella neuroscienza
Le prospettive future nella neuroscienza sono di fondamentale importanza per il progresso della ricerca sulle malattie neurodegenerative e sulla memoria. Dalle ricerche recenti, è emerso che la corteccia entorinale laterale svolge un ruolo cruciale nella formazione e nel richiamo dei ricordi. Tuttavia, sono necessari ulteriori studi per comprendere appieno i meccanismi che regolano questa funzione cerebrale. Inoltre, le metodologie utilizzate per l’individuazione della casa dei ricordi nel cervello stanno continuamente evolvendo, grazie all’uso di tecniche avanzate come la risonanza magnetica funzionale e la stimolazione cerebrale profonda. Queste nuove metodologie aprono nuove possibilità di studio e potrebbero portare a importanti scoperte nel campo delle malattie neurodegenerative.
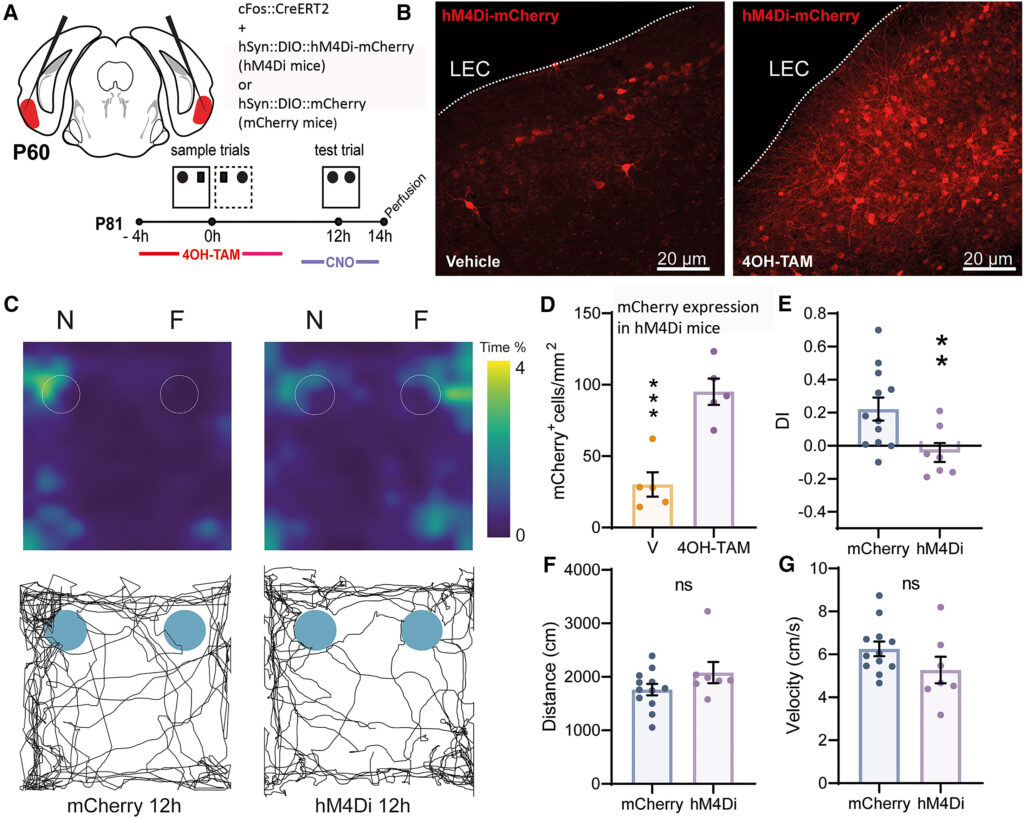
(A) Timeline sperimentale che mostra l'iniezione di due costrutti virali AAV5, cFos::CreERT2 e hSyn::DIO::hM4Di-mCherry, a p60 e il paradigma della memoria OPCRT a p81. Quattro ore prima dell'esecuzione delle prove campione, i topi hanno ricevuto un'iniezione i.p. di 4OH-TAM (25 mg/kg) per indurre la ricombinazione e l'espressione del recettore chemogenetico inibitorio; 30 minuti prima dell'esecuzione della prova, hanno ricevuto un'iniezione i.p. di CNO per indurre l'attivazione del recettore hM4Di.
(B) Immagini rappresentative che mostrano l'espressione di hM4Di-mCherry sia in topi iniettati con il veicolo sia in topi iniettati con 4OH-TAM alle seguenti coordinate: ML = -3,9, AP = -3 e DV = 4,5 da Bregma.
(C) Esempi di heatmap e tracking plot dell'attività esplorativa dei topi (posizione del naso) durante la prova del paradigma OPCRT.
(D) La densità dei neuroni che esprimono hM4Di era significativamente più alta nei topi iniettati con 4OH-TAM rispetto ai topi iniettati con il veicolo (95,15 ± 9 cellule/mm2, n = 5 4OH-TAM vs. 30,16 ± 8 cellule/mm2 veicolo, p = 0,0008, df = 5, t = 5,2, test t a due code non appaiato).
(E) La DI dei topi mCherry 12 ore dopo la presentazione delle prove campione era significativamente più alta del caso (0,22 ± 0,07, n = 12, p = 0,0087, df = 11, t = 3, one-sample t test). Al contrario, la DI dei topi hM4Di non differiva dai livelli casuali (-0,04 ± 0,06, n = 7, p = 0,50). Inoltre, i due gruppi erano significativamente diversi tra loro (mCherry 12 h vs. hM4Di, p = 0,02, df = 17, t = 2,6, test t a due code non accoppiato).
(F) Nessuna differenza significativa nella distanza percorsa durante il test tra i gruppi (1.761 ± 106 cm, n = 12 mCherry vs. 2.079 ± 199 cm, n = 7 hM4Di, p = 0,14, df = 17, t = 1,53, test t a due code non appaiato).
(G) Nessuna differenza significativa nella velocità media tra i gruppi durante la prova (6,26 ± 0,34 cm/s, n = 12 mCherry vs. 5,1 ± 0,56 cm/s, n = 8 hM4Di, p = 0,08, test t a due code non confrontato). ∗∗p < 0,01, ∗∗∗p < 0,001. Barra di scala in (B), 20 μm. I dati sono riportati come media ± SEM.
In conclusione…
In conclusione, le prospettive future nella neuroscienza si concentrano sulla comprensione approfondita dei meccanismi cerebrali coinvolti nella memoria e sullo sviluppo di nuove terapie per trattare le malattie neurodegenerative. La corteccia entorinale laterale riveste un ruolo cruciale nella formazione e richiamo dei ricordi nel cervello umano. Le ricerche recenti hanno permesso di approfondire la comprensione di questa regione cerebrale e delle sue implicazioni nelle malattie neurodegenerative. Le metodologie utilizzate, come l’imaging cerebrale ad alta risoluzione e gli studi su modelli animali, hanno contribuito a delineare le caratteristiche e i meccanismi della memoria. Tuttavia, molte domande rimangono ancora senza risposta. Ad esempio, quali sono le interazioni tra la corteccia entorinale laterale e altre regioni cerebrali coinvolte nella memoria? Come possiamo sfruttare queste conoscenze per sviluppare terapie innovative per le malattie neurodegenerative? Il futuro della neuroscienza sarà caratterizzato da ulteriori scoperte e approfondimenti che ci permetteranno di comprendere appieno il funzionamento del cervello e trovare soluzioni per preservare e ripristinare la memoria.





